
BACKGROUND
IT ALL STARTS IN THE ATMOSPHERE: HOW HEAT, WINDS AND THE ROTATION OF THE EARTH COMBINE TO SHAPE THE OCEAN SURFACE CIRCULATION!
Did you know that the major currents flowing at the surface of the ocean are driven by a combination of wind forcing, the rotation of the Earth - known as Coriolis effect - and the geographical distribution of land and sea?
What happens is that winds are produced in response to inequalities in the distribution of the solar energy flux between the equator and the poles making tropical regions warm and the high-latitude polar regions cold. Then, as winds blow from high- to low-pressure regions, they end up being responsible for the redistribution of heat between low (warm) and high (cold) latitudes. At the same time, these equatorward winds are deflected to the west by the Coriolis effect, giving rise to the NE and SE trade winds in the northern and southern hemisphere, respectively. So, when reaching around 30º to the north and south of the equator, such warm air eventually cools and descends back to the earth’s surface in the subtropical highs, while being transported back to the equator by the trade winds.

When these global gyral-like winds blow over the ocean, they will drive surface currents by dragging the surface of the water and causing it to move and build up towards the direction of the wind, while also being deflected by the Coriolis effect. This is why both wind- and surface ocean global circulation have a similar gyral pattern.
Such circular wind convection cells are known as the Hadley cells, while the zone where the trade winds converge around the equator (between ~5ºN – 5ºS) is the ITCZ, or Intertropical Convergence Zone, a region marked by a permanent low-pressure region loaded with heat and humidity that results in increased convection, cloudiness, and rainfall [1] [2].
SUBTROPICAL GYRES AND EASTERN BOUNDARY CURRENTS
To these major ocean currents forced by the overlying anticyclonic wind systems that occur north and south of the equator (between ~10-15º and ~40-45º) it is given the name of “subtropical ocean gyres”. The poleward-flowing currents on the western sides of the oceans are typically strong and narrow while the currents throughout the remainder of the oceans are generally broader and slow.
Two of the five major ocean-wide gyres in the global ocean are in the Atlantic, i.e., the North Atlantic and the South Atlantic subtropical gyres, each being flanked by a strong and narrow “western current” flowing polewards, and by a weak and broad “eastern current” flowing equatorwards. The North Atlantic gyre is flanked by the very strong western Gulf Stream paired with the eastern Canary Current, while the South Atlantic gyre is flanked by the western Brazil Current paired with the eastern Benguela Current [1][2].
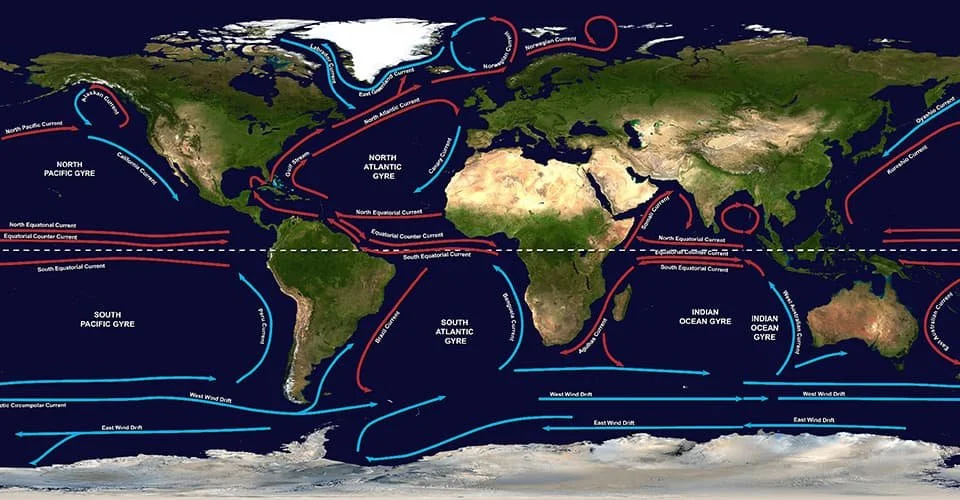
These are the five major oceanic subtropical gyres which are directly linked to the overlying anticyclonic wind systems: the North and South Atlantic Subtropical Gyres, the North and South Pacific Subtropical Gyres, and the Indian Ocean Subtropical Gyre (image from NOAA).
While eastern boundary currents may be considered less impressive compared to the much more striking western currents (e.g., Gulf Stream is ~100 km wide and flows at a surface velocity up to 2 ms-1 fast, while the Canary Current spreads over 1000 km wide, flowing at less than 0.25 ms-1), eastern boundary currents are ecologically important in that they are linked to the occurrence of biologically productive coastal upwelling.

This is a map of the global annual primary production (PP) based on mean photosynthesis versus irradiance (P-I) parameters, for the period of 1998-2018 (image: Kulk et al., 2020).
Upwelling systems are characterized by an upward movement of water parcels in the water column of the ocean over a reasonably long period (several days to weeks). It happens when alongshore wind stress and the Coriolis effect combine to promote the occurrence of Ekman transport towards offshore. Once the upwelled waters reach the sunlight-illuminated upper part of the ocean, the nutrients they carry will fuel primary productivity resulting in phytoplankton to often bloom near the coast. As the upwelled water spreads towards the sea, it will form plume-like pigment-rich filaments which tend to mature while being transported offshore, as seen by an increase in phytoplankton biomass at the expense of the dissolved nutrients, as well as changes in species composition [3].
The intensity of upwelling is not, however, uniform. Rather, it is shaped by temporal variations in wind strength as well as by coastal and seafloor topographic features, such as headlands and submarine canyons. While the upwelling of nutrients usually happens in the period of highest wind stress, it is under the well-stratified conditions during the periods in between that phytoplankton rapidly uptakes nutrients to undertake its highest photosynthetic rate, cell division and population growth while being held in or near the euphotic zone [4].

Ekman transport in the Northern Hemisphere: longshore northerly winds combine with the Coriolis effect to force surface waters to move away from the coast while deeper, cooler and nutrient-rich waters upwell from beneath the thermocline to take their place (image: Merino & Monreal-Gómez, 2011).
WHY ARE EBUS SO IMPORTANT?
Eastern Boundary Upwelling Systems or Ecosystems (EBUS/EBUE) are among the most biologically productive regions of the ocean. Despite of covering only ~1% of the total ocean volume and <3% of the global surface area [5], EBUS are responsible for over 10% of the marine new production [6] supporting roughly 25% of the total worldwide fisheries [7]. Therefore, EBUS provide ecosystem, economic (including fishing and aquaculture), and also recreational services to about millions of people living along their coasts, especially in countries with a strategic vector of development in the economy of the sea.
EBUS are also important for the climate of the Earth, as they play a disproportionately important role in the cycling of carbon and major nutrients in the global ocean with great implications for the regulation of atmospheric CO2. Like land plants, marine phytoplankton need light and nutrients to survive and reproduce in the photic zone of the ocean, taking care of oxygen production and the main responsible for the “biological carbon pump”*. While most of the biological organic material produced in surface waters is recycled in the upper ocean, the biological carbon pump allows for a small yet important fraction of it to be exported and sequestered away from the atmosphere for hundreds to thousands of years. Highly productive regions such as EBUS are, hence, particularly relevant for the global carbon budget, particularly in what concerns the regulation of CO2 levels in the atmosphere.
EBUS AND CLIMATE CHANGE
EBUS are vulnerable to the multiple effects of climate change. All EBUS have experienced extensive fluctuations in pelagic fish stocks and ecological shifts of fish populations that reflect significant food web changes observed over the past 5-6 decades [9]. Long-term variations in abundance, distribution and community structure of plankton are also evident for the same period. However, it is not yet entirely clear, how exactly EBUS are being, and will continue to be affected by climate change.
Studies based on the application of Earth System Models (ESM) predict a reduction in the intensity of wind and upwelling at EBUS located at low latitude regions, and its enhancement at high latitude regions, for RCP 8.5**. On the other hand [10] suggested that increasing land-sea temperature gradients is likely to affect atmospheric pressure cells associated with the development of winds, resulting in intensified wind-induced upwelling and subsequent enhanced NPP within EBUS. Other studies, however, reported inconclusive results that suggest both and increase or a decrease of NPP in response to climate change [11][8]. While coastal warming is expected to render upwelling less effective in lifting nutrient-rich deep waters into the photic zone due to enhanced ocean thermal stratification, an overall uncertainty seems to prevail regarding the future effects of climate change on coastal temperatures and biogeochemistry.
ATLANTIC EBUS: CANARY AND BENGUELA
In PRIMUS we will investigate the characteristics of the Canary Current system (including the Galician Rias and the Portuguese coast, in the Iberian Península) and the Benguela Current system, flowing along the eastern flank of the North and South Atlantic subtropical gyres, respectively.
- Canary Current Upwelling System
The Canary Current upwelling system extends for almost 3700 km from the northern tip of the Iberian Peninsula at 43ºN to the south of Dakar at ~10ºN, influenced by seasonal latitudinal variations of the Azores high-pressure atmospheric system. Upwelling favorable winds occur mainly during summer (and spring) in the northern 1/3, mainly during winter (and fall) in the southern 1/3, and yearlong around the latitude of Cape Blanc (~21º N), offshore Mauritania. About 75% of the fish catch comprises only four types, of which the most abundant are clupeids (sardina pilchardus and sardinella aurita), whose stocks tend to change with seasons and undertaking northward migrations during summer [13][14].
The Galician upwelling region on the NW Iberia is right at the boundary between the North Atlantic subtropical and subpolar regimes, being influenced by the North Atlantic Current and by the Azores Current [15]. An important feature of this region (42º–44ºN) are the Galician rias, a group of large, but relatively narrow, coastal inlets where high productivity and valuable ecosystem services are deeply tied to coastal upwelling in spring and summer [16], producing around 40% of the European, and 15% of the world production of mussels. Any significant change in these patterns is likely to affect both the sustainable exploitation of its abundant marine living resources and their economic value [17], as well as the role of this highly productive region as a natural sink for anthropogenic CO2 [18][15].

Here you get to see the distribution of the mean surface Chl-a concentrations in the Atlantic EBUS during May 2020: the Canary Current System, including the Iberian upwelling region (Galicia and the Portuguese coast) along the NE Atlantic, and the Benguela Current System along the SE Atlantic (image produced and downloaded from OCC ESA).
The Portuguese coast forms a distinct oceanographic and ecological subsystem within the NE Atlantic due to the discontinuity caused by Mediterranean waters flowing from SE. Markedly different shelf systems north and south off Lisbon cause important variations in upwelling-induced nutrient availability in the euphotic zone, and in the water column stability [19][29][21][22]. Further north, the shelf is wider and more heavily subjected to rainfall and freshwater river discharge, while the southern shelf is narrower, less subjected to freshwater input, and marked by more headlands and other coastal irregularities. As a result, phytoplankton blooms earlier off the north coast because of the stratification caused by the buoyancy of the freshwater in the winter-spring transition. Stronger and more persistent winds in this region also contribute more nutrient-rich upwelled water giving rise to higher NPP and zooplankton biomass. By contrast, lower freshwater input off the south coast forces marine phytoplankton to wait for the surface water to become warm enough to produce the required stratification conditions driving the spring bloom. In addition, the narrower southern shelf makes it faster for the upwelled water to cross it, which translates into will less time for zooplankton to develop at the expense of phytoplankton. This results in smaller zooplankton populations in the south, with much of the ungrazed phytoplankton sinking to the bottom to support a higher proportion of benthic fish. Greater influence of subtropical waters in the southern shelf also leads to a greater species diversity in the plankton south of Lisbon [23][4].
The NW African coast is an important part of the Canary Current upwelling system, hosting high productivity and fisheries which are essential for the livelihood of several African countries (e.g., Morocco, Mauritania, Senegal, Cape Verde, and nearby African countries) as well as the Canary Islands. At the latitude of Cape Blanc (~21°N), the Canary Current detaches from the continental shelf and veers towards southwest direction, feeding into the North Equatorial Current. There, the southward flowing Canary Current meets the northward flowing Mauritanian Current and the so-called Cape Verde Frontal System (CVFS) is formed [24], separating colder subtropical- from warmer tropical waters. The CVFS is a region of significant offshore water export [25][26] favoring the advection of cold- and nutrient- and filament-rich surface water driven by yearlong coastal upwelling off Cape Blanc. The so-called “Giant Cape Blanc Filament” [27][28] is a permanent upwelling filament extending up to 600 km offshore where it interacts with the deep-ocean boundary [28]. Due to limited local infrastructure, this region has been relatively poorly sampled compared to European upwelling regions, despite of covering a large area of the northern and western coast of Africa.
- Benguela Current Upwelling System
The Benguela upwelling system is related to the equatorward-flowing Benguela Current, which spreads over ~200 km wide along the coastlines of Angola, Namibia and South Africa, extending from Cape Town north to ~15º S, being influenced by seasonal latitudinal variations of the South Atlantic high-pressure atmospheric system. Since the Benguela system is in the southern hemisphere, the offshore Ekman transport of water occurs in response to southerly winds blowing parallel to the coast. Upwelling near Cape Town is usually stronger during summer, while the very major upwelling at Cape Frio is strongest during winter. Like the Canary Current system, optimal conditions for PP along the Benguela Current system occur in response to an alternation between the upwelling phase, during which surface waters become nutrient-replenished, and the quiescent phase that follows, during which stratification establishes and phytoplankton biomass increases. A lag of 1-4 days occur between the nutrient input and the phytoplankton bloom, the latter normally lasting 4-10 days [4].

As in other upwelling systems worldwide, sardines are amongst the most abundant fishes benefiting from highly productive upwelling conditions in the Canary and Benguela upwelling systems (Image credits here).
High primary productivity in this region supports large scale commercial fishing enterprises, small-scale subsistence fishing endeavours and marine aquaculture. As in other upwelling systems worldwide, sardines and anchovies are the most abundant fishes, with the stocks being roughly divided into those breeding in the Cape area, and those breeding north off Namibia. Fish species targeted by various role players in the Benguela Current system are, at some point in their life cycle, dependant on the availability of plankton for their successful growth and recruitment. Marine primary production also plays an important role in biogeochemical cycling (air-sea carbon exchange processes) within the region [29].

References
[1] Ross, D., 1995. Introduction to oceanography. Pub. New York, NY : HarperCollins College Publishers, 496 pp.
[2] Colling, A. 2001. Ocean Circulation. Edited by The Open University, ISBN 0750652780, 288 pp.
[3] Mohrholz, V. et al., 2014. Cross shelf hydrographic and hydrochemical conditions and their short term variability at the northern Benguela during a normal upwelling season, Journal of Marine Systems, 140, 92-110. https://doi.org/10.1016/j.jmarsys.2014.04.019
[4] Mann, K. H. and Lazier, J. R., 2006. Dynamics of Marine Ecosystems, Biological-Physical Interactions in the Oceans. 3 Edn., Blackwell Publishing, ISBN: 978-1-405-11118-8, Malden, MA, Oxford, UK, 512 pp.
[5] Bonino, G. et al., 2019. Interannual to decadal variability within and across the major eastern Boundary Upwelling Systems. Scientific Reports 9, 19949. https://doi.org/10.1038/s41598-019-56514-8
[6] Beca-Carretero, P.P. et al., 2019. Seasonal and inter-annual variability of net primary production in the NW Iberian margin (1998–2016) in relation to wind stress and sea surface temperature. Progress in Oceanography, 178, 102135. https://doi.org/10.1016/j.pocean.2019.102135
[7] Messié, M., Chavez, F.P., 2015. Seasonal regulation of primary production in eastern boundary upwelling systems. Prog. Oceanogr. 134, 1–18. https://doi.org/10.1016/j.pocean.2014.10.011
[8] García-Reyes, M. et al., 2015. Under Pressure: Climate Change, Upwelling, and Eastern Boundary Upwelling Ecosystems. Frontiers in Marine Science 2, 109. https://doi.org/10.3389/fmars.2015.00109
[9] Verheye, H.M. et al., 2016. Plankton productivity of the Benguela Current Large Marine Ecosystem (BCLME). Environmental Development, 17, 75-92. http://dx.doi.org/10.1016/j.envdev.2015.07.011
[10] Bakun, A., 1990. Global climate change and intensification of coastal ocean upwelling. Science 247, 198–201. http://dx.doi.org/10.1126/science.247.4939.198
[11] Abrahams, A. et al., 2021. Variation and Change of Upwelling Dynamics Detected in the World’s Eastern Boundary Upwelling Systems. Frontiers in Marine Science, 8, 626411. https://doi.org/10.3389/fmars.2021.626411
[12] Cermeño, P. et al., 2008. The role of nutricline depth in regulating the ocean carbon cycle. PNAS 105 (51), 20344–20349. https://doi.org/10.1073/pnas.0811302106
[13] Trumble, R.J. et al., 1981. Seasonal food production and consumption by nekton in the northwest African upwelling system. In Coastal Upwelling (ed. F.A. Richards) 458-463, Am. Geophys. Union, Washington, DC.
[14] Palomera, I., Rubies, P., 1982. Kinds and distribution of fish eggs and larvae off northwest Africa in April/May 1073. Rapp. P. Reún. Const. Int. Explor. Mer. 180, 356-358.
[15] Álvarez-Salgado, X.A. et al., 2010. Iberian margin: the Rías. In: Liu, K.-K., Atkinson, L., Quiñones, R., Talaue–McManus, L. (Eds.), Carbon and Nutrient Fluxes in Continental Margins: A Global Synthesis, IGBP Book Series. Springer, Berlin, pp. 103–120.
[16] Arístegui, J. et al., 2009. Sub-regional ecosystem variability in the canary current upwelling. Prog. Oceanogr. 83, 33–48. https://doi.org/10.1016/j.pocean.2009.07.031
[17] Alvarez-Salgado, X.A. et al., 2008. Renewal time and the impact of harmful algal blooms on the extensive mussel raft culture of the Iberian coastal upwelling system (SW Europe). Harmful Algae 7 (2008) 849–855. https://doi.org/10.1016/j.hal.2008.04.007
[18] Gago, J. et al., 2003. Assessing the fate of dissolved and suspended organic carbon in a coastal upwelling system (Ría de Vigo, NW Iberian Peninsula). Estuar. Coast. Shelf Sci. 56, 271–279. https://doi.org/10.1016/S0272-7714(02)00186-5
[19] Moita, M.T. et al., 2003. Distribution of chlorophyll a and Gymnodinium catenatum associated with coastal upwelling plumes off central Portugal. Acta Oecologica 24, S125–S132. https://doi.org/10.1016/S1146-609X(03)00011-0
[20] Silva A. et al., 2009. Composition and interannual variability of phytoplankton in a coastal upwelling region (Lisbon Bay, Portugal). Journal of Sea Research 62, 238-249. https://doi.org/10.1016/j.seares.2009.05.001
[21] Guerreiro, C. et al., 2013. Late winter coccolithophore bloom off central Portugal in response to river discharge and upwelling. Continental Shelf Research 59, 65 83. http://dx.doi.org/10.1016/j.csr.2013.04.016.
[22] Oliveira P.B. et al., 2019. Phytoplankton distribution and physical processes off NW Iberia during two consecutive upwelling seasons, Continental Shelf Research, 190. https://doi.org/10.1016/j.csr.2019.103987
[23] Cunha, M.E., 2002. Physical control of biological processes in a coastal upwelling system: comparison of the effects of coastal topography, river runoff and physical oceanography in the northern and southern parts of Portuguese coastal waters. Unpublished PhD thesis, University of Lisbon, Portugal.
[24] Pastor, M.V. et al., 2012. Meridional changes in water mass distributions off NW Africa during November 2007/2008. Cienc. Mar. 38 (1B), 223–244. https://doi.org/10.7773/cm.v38i1B.1831
[25] Pastor, M.V. et al., 2008. Water and nutrient fluxes off northwest Africa. Cont. Shelf Res. 28, 915–936. https://doi.org/10.1016/j.csr.2008.01.011
[26] Alonso-González, I.J. et al., 2009. Lateral POC transport and consumption in surface and deep waters oft he Canary Current region: a box model study. Glob. Biogeochem. Cycles 23, GB2007. https://doi.org/10.1029/2008GB003185
[27] Van Camp, L. et al., 1991. Upwelling andboundary circulation off Northwest Africa as depicted by infrared and visible satellite observations. Prog. Oceanogr. 26, 357–402. https://doi.org/10.1016/0079-6611(91)90012-B
[28] Gabric, A.J. et al., 1993. Offshore export of shelf production in the Cape Blanc (Mauritania) giant filament as derived from coastal zone color scanner imagery. J. Geophys. Res. 98 (C3), 4697–4712. https://doi.org/10.1029/92JC01714
[29] Silió-Calzada, A. et al., 2008. Estimation of new primary production in the Benguela upwelling area, using ENVISAT satellite data and a model dependent on the phytoplankton community size structure. Journal of Geophysical Research, 113, C11023.https://doi.org/10.1029/2007JC004588